|
1.3.2. Components of the Carbon Balance of Ecosystems
To understand the fluxes of carbon between a specific ecosystem and the atmosphere
over a specified period of time and to appreciate its sensitivity to current
environmental conditions, disturbance, and climate change, we need to consider
the principal component processes within the system that add up to give the
overall NEP. The feedbacks among these processes, their speed of response, and
their sensitivity to environmental change determine NBP-the future and long-term
carbon sequestration potential of the ecosystem.
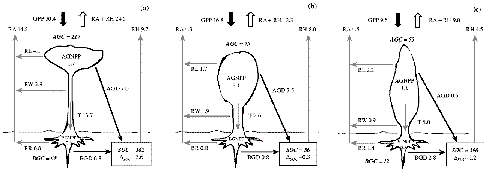
Figure 1-3 shows measured annual carbon fluxes in
a typical boreal, temperate, and tropical forest. NEP is the difference between
the gross input of carbon in photosynthesis, GPP, and the sum of the losses
of carbon in autotrophic respiration (RA) and RH. The component fluxes accumulated
over a year should add up to the annual NEP, which is measured independently
as described below (see Section 1.3.2.3). A mass balance
of the component fluxes above ground (photosynthesis; foliage, branch, and stem
respiration; leaf, branch, and stem litter production; and aboveground NPP)
enables an estimate to be made of the amount of carbon internally translocated
below ground. A mass balance of the component fluxes below ground (the inputs
of litter and translocate from above, root and heterotrophic respiration, fine
root turnover, mycorrhizal and root system NPP) enables an approximate estimate
to be made of net changes in the pool of soil carbon. There are appreciable
errors, however, in measuring all component fluxes (below-ground fluxes in particular),
so close agreement between the two estimates of NEP is not to be expected.
The processes of photosynthesis and respiration are functions of several environmental
and plant variables, including solar radiation, air and soil temperature and
humidity, availability of water and nutrients, atmospheric ozone and other pollutants,
leaf area, and foliar nutrition. Climate change therefore affects these processes
in several ways. Photosynthesis is likely to be reduced by an increase in cloud
cover but increased by enhanced global atmospheric CO2 concentration and, on
some sites, by atmospheric nutrient deposition. All respiratory processes are
sensitive to temperature, as is the rate of population growth of respiring organs-particularly
the fine roots and heterotrophic organisms in the soil. Thus, "soil respiration"
is a function of soil temperature (e.g., Boone et al., 1998; Rayment
and Jarvis, 2000), which, if increased, leads in the short term to enhanced
mineralization of soil organic matter and the release of nutrients-which, in
turn, feed back to stimulate photosynthesis, increased leaf area, and tree growth.
Evidence is accumulating, however, that in the longer term soil respiration
acclimates to the rise in temperature and stabilizes at close to the original
rate. In semi-arid and arid regions in particular, the availability of soil
water-and thus changes in rainfall patterns-are also of utmost importance for
the balance between carbon gains and losses.
|
|
Figure 1-3: Estimated annual total carbon stocks
and flows for three representative forest stands in tropical, temperate,
and boreal regions. Stocks in bold italics are in tons of carbon per
hectare (t C ha-1). Flows are in t C ha-1 yr-1. (a) Tropical rain forest
near Manaus, Amazonia, Brazil; (b) temporate deciduous oak-hickory forest,
near Oak Ridge, Tennessee, USA; and (c) boreal evergreen black spruce
forest, near Prince Albert, Saskatchewan, Canada.
|
|