3.3.3. Terrestrial Ecological Systems
Climate change will affect terrestrial ecological systems through changes in
permafrost as well as direct climatic changes, including changes in precipitation,
snow cover, and temperature. Terrestrial ecoystems are likely to change from
tundra to boreal forests, although vegetative changes are likely to lag climatic
change. Vegetational ecosystem models suggest that the tundra will decrease
by as much as one- to two-thirds of its present size (see Annex C). Climate change is expected to occur at a rapid rate relative to the
speed at which forest species grow, reproduce, and reestablish themselves or
to their ability to develop appropriate soils. The boreal forest covers approximately
17% of the world's land surface in a circumpolar complex of forested and partially
forested ecosystems in northern Eurasia and North America (IPCC 1996, WG II,
Section 1.6.1). If there is warming, the greatest forest changes are expected
in high latitudes. Polar projections of this region's terrestrial ecological
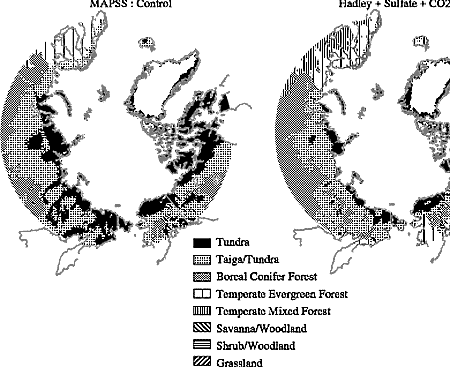
changes (Figure 3-3) derived from two different models'
depictions of the future climate reinforce this conclusion, showing significant
reduction in the tundra and taiga biomes and expansion of the boreal forest.
 |
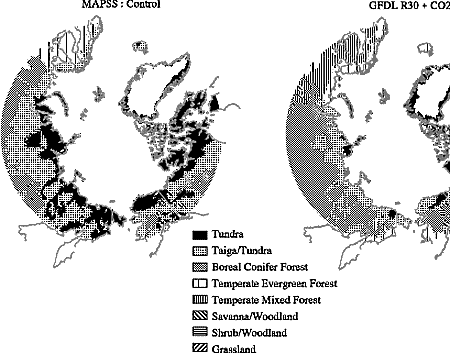 |
| Figure 3-3: Ecological shifts derived from the MAPSS model in response
to changes projected by two GCM simulations of future climate. |
A poleward migration of the northern tree line would decrease winter albedo
because the tree canopy has much lower albedo than exposed snow surfaces, affecting
local climate through increased absorption of the sun's incoming energy (IPCC
1996, WG II, Chapter 6). Large losses are projected in the area of boreal forests
despite their encroachment into current tundra. Shrinkage in total area because
of the geographically limited poleward shift leads to a net loss of about 25%
in boreal coverage (IPCC 1996, WG II, Section 1.3.4). On the other hand, vegetative
ecosystem modeling suggests an expansion in the boreal forest ranging from 108%
to 133% of the present size (see Annex C). The species
composition of forests is likely to change; entire forest types may disappear,
while new assemblages of species, and hence new ecosystems, may be established
(IPCC 1996, WG II, Section 3.1). The rate of migration is critical, and it is
likely that the loss from the southern margin will be greater than the gain
in the northern margin. Projections for habitat changes are presented in Table
3-1.
| Table 3-1: Area of each biome
(in 1,000 km2) north of the Arctic Circle, under current and future climate(s),
as projected by three models discussed in Annex C.
|
|
| |
|
MAPSS + CO2 Effect |
|
| |
Present |
OSU |
GFDLR30 |
UKMOS |
| Ice (perennial land-mass ice) |
1,745 |
1,437 |
1,328 |
1,532 |
| Tundra |
4,316 |
2,972 |
2,019 |
1,832 |
| Taiga/Tundra |
1,950 |
2,520 |
2,632 |
2,265 |
| Boreal Conifer Forest |
0 |
791 |
1,432 |
1,643 |
| Other |
178 |
468 |
778 |
918 |
By 1995, the Arctic contained 285 protected areas covering 2.1 million km2
(UNEP, 1997). There will be considerable impacts of climate change on resource
management in the tundra (IPCC 1996, WG II, Section 7.5). There is likely to
be a change in the migration patterns of polar bears and numbers of caribou,
along with other biological impacts (IPCC 1996, WG II, Section 7.5). High-arctic
Peary caribou and musk-oxen may become extinct, and barren-ground caribou and
musk-oxen elsewhere may be reduced (Gunn, 1995). Arctic island caribou migrate
seasonally across the sea ice between many of the arctic islands in late spring
and fall; changes in sea ice would disrupt those migrations, with unforeseen
consequences to population survival and gene flow.
It is also anticipated that small mammals, from lemmings to aquatic furbearers
(muskrats, beavers, mink), will face ecosystem alterations that will change
their abundance and distribution. Amphibians and reptiles can be expected to
increase their range, and the distribution and abundance of hundreds of arthropod
species will alter. Changes in the timing and abundance of forage availability
and parasite infestations may accumulate-driving populations into decline, with
serious consequences for people still depending on them (Jefferies, 1992). Increased
incidences of forest fires and insect outbreaks are likely, such as already
observed in Alaska.
Indirect temperature effects associated with changes in tundra thaw depth,
nutrient availability, and vegetation will cause substantial changes in Arctic
species composition, litter quality, and nutrient availability. Increased nutrient
availability increases shrub abundance and decreases the abundance of mosses,
an important soil insulator (IPCC 1996, WG II, Section 2.7.3).
Some alpine and tundra plants have no photoperiod requirements for initiating
growth and therefore are affected by spring frosts. For example, in northern
Sweden, increased length of the growing season and increased summer temperature
will have a significant impact on reproduction and population dynamics in Arctic
and alpine plants (IPCC 1996, WG II, Section 5.2.3.1). Experimental doubling
of CO2 concentration has relatively little multiyear effect on plant growth
in Arctic tundra, presumably because of constraints of low nutrient supply (IPCC
1996, WG II, Section 2.2.2).
Major shifts in biomes will be associated with changes in microbiological (bacteria,
algae, etc.) and insect communities. As a consequence, some may diminish while
others prosper. All microbiologically facilitated processes are strongly affected
by moisture and temperature. Annual soil respiration rates are likely to increase
because of the lengthened season for breakdown of plant material and because
increasing temperature strongly stimulates organic-matter decomposition, especially
in Arctic regions subject to permafrost (IPCC 1996, WG II, Section A.3.1).
(continues on next page...)
|