5.7.3.1. Physical Conditions
5.7.3.1.1. Ice cover
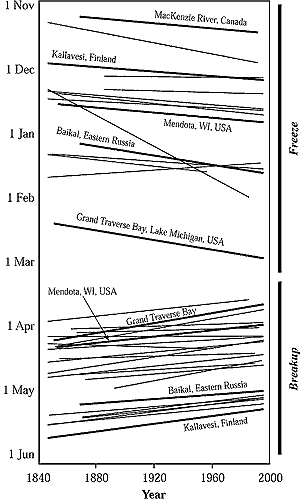
Figure 5-6: Historical trends in freeze and breakup dates of lakes
and rivers in the Northern Hemisphere; 37 of the 39 trend slopes are in
the direction of warming (modified from Magnuson et al., 2000). |
To date, the only limnological properties measured or simulated
at a global scale are lake and river ice phenologies. Ice-cover durations for
inland waters have decreased over the entire northern hemisphere (Magnuson et
al., 2000) (see Figure 5-6). Change from 1846 to 1995 averaged
8.7 days later freeze and 9.8 days earlier break-up; these changes correspond
to a 1.8°C increase in air temperature. Lake Suwa in Japan, Tornionoki River
in Finland, and Angara River in Siberia have longer records; ice phenologies
have been changing in the direction of warming since about the early 1700s,
but at slower rates than during 1846-1995. Interannual variability in freeze
dates, thaw dates, and ice duration are increasing (Kratz et al., 2001). Interannual
variability was greater from 1971 to 1990 than from 1951 to 1970 in 184 lakes
in the northern hemisphere.
For the Baltic Sea, which contains freshwater communities near shore and to
the north, simulation until 2050 reduced the extent of ice cover from 38% of
the area to 10%, and simulation until 2100 reduced ice cover to zero (Haapala
and Leppäranta, 1997). For U.S. lakes, ice-on date, ice-off date, ice duration,
ice thickness, and the continuity of ice cover all responded to a 2xCO2
change (Fang and Stefan, 1998). Mean duration at 209 locations distributed evenly
across the country declined by 45 days (60%), and ice thickness by 21 cm (62%).
Simulated profiles of deepwater oxygen concentrations and water temperatures
also changed dramatically in lakes across the United States (Fang and Stefan,
1997).
Lake ice influences biogeochemical cycling, including gas exchange with the
atmosphere, fish habitat availability (through changes in pH and dissolved oxygen),
biodiversity, and seasonal succession (Arnell et al., 1996; Cushing, 1997).
Decreased ice cover should reduce winterkill of fish.
Ice break-up date influences the productivity of diatoms beneath the ice of
Lake Baikal (Granin et al., 2001). Seasonal succession of phytoplankton in small
lakes is altered partly because of variation in the incoculum of algae beneath
the ice in different years (Adrian and Hintze, 2001). The mechanism for enhanced
diatom productivity in Lake Baikal is a combination of nutrient enrichment immediately
beneath the ice owing to the extrusion of salts as the water freezes and vertical
mixing that results from the greater density of the water with extruded salts
immediately beneath the ice. Reductions in ice thickness and increases in snow
would be expected to reduce diatom production.
River ice plays an important biological role by regulating flow aeration and
oxygen concentrations under the ice (Prowse, 1994; Chambers et al., 1997); dissolved
oxygen can approach critical levels for river biota (e.g., Power et al., 1993).
Decreases in the duration of the river-ice season or increases in the size and
frequency of open-water sections where re-aeration can occur will reduce anoxia.
Warmer winters favor the formation of mid-winter break-ups produced by rapid
snowmelt runoff, particularly those initiated by rain-on-snow events. Such events
significantly impact benthic invertebrate and fish populations where late-season
break-ups are the norm (Cunjak et al., 1998).
Ecological impacts will be influenced most by changes in break-up timing and
intensity. Physical disturbances from break-up scouring and flooding influence
nutrient and organic matter dynamics, spring water chemistry, and the abundance
and diversity of river biota (Scrimgeour et al., 1994; Cunjak et al., 1998).
Ice-induced flooding supplies the flux of sediment, nutrients, and water that
are essential to the health of freshwater delta ecosystems (e.g., Lesack et
al., 1991; Prowse and Conly, 1998). Even the mesoscale climate of delta ecosystems
depends on the timing and severity of break-up flooding (Prowse and Gridley,
1993).
Climate-induced change in ice cover or the timing and severity of its break-up
will affect the movement and deposition of sediment and associated contaminants.
Stable cover leads to deposition of sediment particles that would remain in
suspension without ice cover. Environmental contaminants with an affinity for
adsorption to fine-grained sediments are more likely to be deposited during
ice-covered, low-flow season than during open-water season. The break-up plume
contains the winter-long accumulation of contaminated sediment (Milburn and
Prowse, 1996). Sediment deposition zones often are where biological productivity
is greatest, such as on river deltas (Milburn and Prowse, 1998). Because growth
rates of aquatic organisms in cold regions are low and because higher trophic
level animals are longer lived, potential exists for greater lifelong accumulation
of contaminants in these aquatic ecosystems than in more temperate regions.
River ice is a key agent of geomorphologic change; it is responsible for creating
numerous erosional and depositional features within river channels and on channel
floodplains (e.g., Prowse and Gridley, 1993; Prowse, 1994). During break-up,
these processes cause channel enlargement and removal and succession of riparian
vegetation. Climatic conditions that alter the severity of such events will
greatly influence river morphology and riparian vegetation.
|