5.2.1. Observational Studies
|
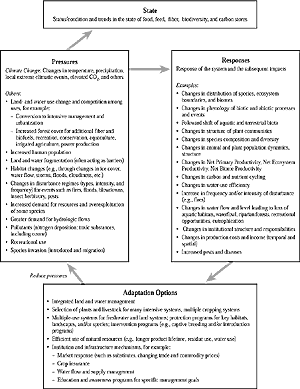
Figure 5-1: Generalized diagram of the state of
specific goods and services that ecosystems provide; how these goods
and services are affected by the multiple pressures of climate change
and human activities; and how the system responds (autonomous adaptation
as in Chapter 19), thus affecting the provision
of goods and services. Adaptation options reduce the impacts and thus
change the vulnerability of the system.
|
There is now a substantial number of observational and
experimental studies that demonstrate the link between climate and biological
or physical processes in ecosystems. The authors of this chapter assembled a
database of more than 2,500 studies that address climate and either a physical
process (e.g., melting of ice on lakes) or a biological factor (e.g., spring
arrival time) of an animal or plant. Most of these studies address experiments
that are valuable primarily in helping to understand the biological mechanisms
prompting the responses of plants and animals to climate but are not helpful
in detecting patterns of change. Many of these studies were conducted over a
period of shorter than 10 years. Because at least 10 years of data are needed
to show a possible trend, this narrowed the number of studies to approximately
500. Because temperature is the variable that can most reliably be predicted
with increasing GHGs, only studies that addressed temperature as the climatic
variable were examined, leaving approximately 250 studies. All 250 studies then
were examined to determine if they met at least two of the following criteria:
- The authors found a statistically significant correlation between temperature
and a species trait (e.g., egg-laying date, location of range boundaries)
or physical process.
- The authors found a statistically significant change in the species trait
over time.
- The authors found a statistically significant change in temperature over
time.
In some cases, the criteria were met by two companion papers rather than a
single paper. These criteria narrowed the qualifying studies to 60; seven were
companion papers. Among the 60 studies, 16 look at physical processes, 10 examine
vegetation changes, eight look at invertebrates, six investigate amphibians
and reptiles, 26 examine birds, and one addresses mammals. Some of these studies
investigate multiple taxa (e.g., bird and insect) in the same paper. A total
of 39 physical processes, 117 plants, 65 insects, 63 amphibians and reptiles,
209 birds, and 10 mammal species were examined in the 43 studies (summarized
in Table 5-3). Approximately 39% of these species
showed no change. Changes in the other 61% included earlier ice-off and later
freeze dates in inland lakes and streams, earlier breeding times, shifting to
higher elevations or latitudes, and changes in densities, development, morphologies,
and genetics.
Several lines of evidence indicate lengthening of the vegetative growing season
by 1.2-3.6 days per decade in the Northern Hemisphere, particularly at
higher latitudes where temperature rise also has been greatest. This lengthening
involved earlier onset of spring and later onset of fall. Summer photosynthetic
activity [based on Normalized Differential Vegetation Index (NDVI) estimates
from satellite data] increased from 1981 to 1991 (Myneni et al., 1997),
concurrent with an advance (by 7 days) and an increase in amplitude of the annual
CO2 cycle since the 1960s, most intensely during the 1980s (Keeling
et al., 1996). Phenological/climate models for Finland indicate an overall
increase in growing season length since 1900 (Carter, 1998). These physical
measures are in accord with observations on organisms. In controlled, mixed-species
gardens across Europe, a lengthening of the growing season by 10.8 days occurred
from 1959 to 1993 (Menzel and Fabian, 1999). Likewise, a study of 36 species
in the central United States documented advances in flowering dates by an average
of 7.3 days from 1936 to 1998 (Bradley et al., 1999).
Responses to increased atmospheric CO2 have been detected in increased
stomatal densities in the leaves of temperate woodland plants (Beerling and
Kelly, 1997). Recent changes (over 9- to 30-year periods) in community composition
have occurred at protected sites in the lower United States and Alaska, concurrent
with local warming trends (Chapin et al., 1995; Brown et al.,
1997a; Alward et al., 1999). Results of warming experiments coupled with
previous knowledge of species' habitat requirements implicate climate as
one factor in these community reorganizations, but additional effects of multiple
pressures have led to complex responses that were not always predicted by bioclimatic
theory (Schneider and Root, 1996).
Multiple studies of treelines at high latitudes in the northern hemisphere
have shown 20th century poleward shifts, often measured as increased growth
at northern boundaries and decreased growth at southern boundaries. Interpretation
of these trends is not straightforward because most change occurred during the
early 20th century warming and the trends have been less pronounced or absent
in recent warm decades (Kullman, 1986, 1990; Hamburg and Cogbill, 1988; Innes,
1991; Lescop-Sinclair and Payette, 1995; Jacoby and D'Arrigo, 1995; Briffa
et al., 1998). These authors have hypothesized that the general lack
of response to recent warming is a result of increases in water stress, severity
of insect attack, and UV radiation and trends toward earlier snowmelt or to
sunlight becoming a limiting growth factor. In addition, some localities that
showed warming and increased growth in the early 20th century have shown cooling
and stable growth since the1970s (Kullman, 1991, 1993). In contrast, simple
predictions of range shifts have been fulfilled in alpine herbs, which have
moved to higher altitudes concurrent with warming in Switzerland (Grabherr et
al., 1994), and loss of low-elevation pine forests in Florida as sea-level
rise has caused toxic levels of salination near coastal areas (Ross et al.,
1994).
| Table 5-3: The number of species and processes
in each region that were found in each particular study to be significantly
associated with regional temperature change. For inclusion in the table,
each study had to meet two of the following three criteria: species or processes
changing over time; regional temperature changing over time; and significant
association between how the temperature and species or processes were changing.
The first number indicates the number of species or processes changing in
the manner predicted with global warming. The second number is the number
of species or processes changing in a manner opposite to that predicted
with a warming planet. When considering those species that have shown
a change, 80% are changing in the manner expected with global warming, while
20% are changing in the opposite direction. Note that about 61 of all
species examined did not show a statistically significant change. References
for each cell are located below the table and collated by row number and
column number (e.g., references for European birds are under E,5—row
E, column 5). "—" indicates that no studies were found for
this region and category. |
 |
| Region |
Column 1:
Lake and
Stream Ice
|
Column 2:
Vegetation
|
Column 3:
Invertebrate
|
Column 4:
Amphibians
and Reptiles
|
Column 5:
Birds
|
Column 6:
Mammals
|
|
| Row A: Africa |
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
| Row B: Antarctica |
—
|
—
|
2
|
0
|
—
|
—
|
—
|
—
|
2
|
0
|
—
|
—
|
| Row C: Asia |
3
|
0
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
| Row D: Australia |
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
—
|
| Row E: Europe |
8
|
0
|
13
|
1
|
46
|
1
|
7
|
0
|
258
|
92
|
7
|
0
|
| Row F: North America |
18
|
0
|
32
|
11
|
—
|
—
|
—
|
—
|
17
|
4
|
3
|
0
|
| Row G: Latin America |
—
|
—
|
—
|
—
|
—
|
—
|
22
|
0
|
15
|
0
|
—
|
—
|
| Total |
29
|
0
|
47
|
12
|
46
|
1
|
29
|
0
|
292
|
96
|
10
|
0
|
|
|